Бактериальная клетка является полноценным живым организмом. Чтобы понимать, как происходит организация обменных процессов в ней, необходимо знать химический .
Химический состав бактериальной клетки для построения белков, углеводов и липидов включает такие элементы: азот, водород, кислород, углерод. Если брать количественное соотношение, то процент воды достигает 85, а сухое вещество, состоящее из белков, минеральных соединений, липидов, углеводов, аминокислот, составляет оставшиеся проценты.
Морфология клеточной стенки
Клеточная стенка ─ это наружный слой, защищающий клетку от внешних воздействий и придающий ей постоянную форму, к примеру, округлую или форму вибриона, палочки. Клеточная стенка играет функцию скелета. Она такая же плотная, как и растительная оболочка, в отличие от животной клеточной оболочки, которая мягкая. Внутри самой клетки давление в несколько раз больше, чем атмосферное, и если бы не ее плотная оболочка, то ее разорвало бы. Толщина стенки варьируется от 0,01 до 0,04 мкм и составляет в среднем 10-50% от общей массы.
Основным химическим компонентом всех изученных ранее видов есть муреин. Он состоит из аминосахаров и аминокислот.
В зависимости от того, как реагирует на окрашивание по Граму, микроорганизмы делятся на грамположительные и грамотрицательные. У грамположительных видов стенка может окрашиваться анилиновыми красителями и после обработки йодом и спиртом сохраняет цвет. Грамотрицательные после реакции на спирт становятся бесцветными. У грамположительных и грамотрицательных видов химический состав бактериальной клетки отличается.
В составе стенки грамположительных форм есть мукопептиды, полисахариды, тейхоевые кислоты.
Клеточная стенка грамотрицательных бактерий по своему химическому составу несколько сложнее. В ней есть большое содержание липидов, которые вместе с белками и сахарами образуют сложные соединения: липополисахариды и липопротеиды. Клеточные стенки грамотрицательных бактерий муреина содержат меньшее количество, но сама структура стенки имеет более сложное строение, чем у грамположительных.
Она состоит из нескольких слоев:
- внутреннего, состоящего из муреина;
- за ним слой, состоящий из белков;
- после идет слой липополисахаридов;
- внешний, состоящий из липопротеидов.
Клеточная стенка пропускает внутрь необходимые вещества, из нее выходят продукты жизнедеятельности.
Капсула
У большинства бактерий над клеточной стенкой есть капсула, которая не относится к обязательным компонентам. Поперечный разрез капсулы может быть в несколько раз больше размеров самой клетки. Микрокапсула имеет ультратонкую структуру, рассмотреть ее можно только при хорошем увеличении. Чаще всего капсула образуется при возникновении неблагоприятных условий для жизни. Капсула выполняет защитные функции. Она защищает клетку от высыхания и принимает непосредственное участие в водном обмене. Состав включает в себя в основном полисахариды. Иногда в капсуле могут находиться клетчатка, полипептиды, гликопротеины.
Морфология цитоплазмы
Все внутренние органоиды находятся в жидкой среде ─ цитоплазме. У нее сложная структура, которую стало возможным изучить лишь с появлением мощных микроскопов. Внешний слой цитоплазмы называется цитоплазматической мембраной. Сквозь мембрану происходит поступление необходимых веществ и выход продуктов жизнедеятельности за пределы клетки. Через мембрану необходимые вещества проникают благодаря сложным биохимическим процессам. В мембране организация нахождения ферментов имеет упорядоченный характер, что позволяет мембране выполнять функции контроля, не давая возможности одним ферментам разрушать другие. Химический состав мембраны состоит из липопротеинов. Мембрана является очень прочной, и с ее помощью клетка может некоторое время жить без оболочки. На тонких срезах под микроскопом хорошо видно, что структура мембраны состоит из слоя липидов, который находится между слоями белков.
Рибосомы
В цитоплазме находятся частицы, осуществляющие синтезирование белка, ─ это рибосомы. Их состав включает белок и молекулы РНК. Число рибосом может насчитываться более тысячи. У многих микроорганизмов рибосомы находятся в свободном состоянии внутри цитоплазмы, но есть виды, у которых рибосомы связаны с мембраной.
Внутри цитоплазмы есть и временные включения, которые являются запасным вариантом и образуются, когда наступают неблагоприятные условия. Такие включения служат дополнительным источником пополнения углерода.
У многих видов микроорганизмов в цитоплазме есть гранулы, в химический состав которых входит крахмал или полисахариды, могут встречаться и капельки жира. Так, например, волютин является довольно распространенным клеточным включением. Его гранулы состоят из полиметафосфата. Обычно микробы аккумулируют волютин в несвойственных для них источниках питания, например, там, где нет серы. Жидкая среда цитоплазмы содержит:
- белки;
- пигменты;
- сахара;
- ферменты;
- аминокислоты.
Жгутики
Жгутики являются дополнительными органоидами бактерий и служат для передвижения клетки. Жгутики есть не у всех микроорганизмов. Крепятся жгутики к цитоплазматической мембране с помощью пары дисков. Количество жгутиков может быть разным: от пары, располагающейся с одного конца, до жгутиков, располагающихся по всей поверхности. Длина жгутиков бактерий может быть в несколько раз больше длины самой клетки. Химический состав жгутиков представлен белковым веществом флагеллином.
Споры
При возникновении негативных химических или физических воздействий бактерии образуют споры, в отличие от грибов, которым споры необходимы для размножения. Для обнаружения спор бактерий используется метод нахождения по Ожешко. Он позволяет выявлять споры благодаря наличию в оболочке спор кальциевой соли. Размеры спор у разных бактерий отличаются, что немаловажно при их дифференциации. Жизненный цикл спор включает в себя такие этапы:
- подготовительный;
- стадия предспоры;
- создание оболочки;
- стадия созревания споры;
- стадия спокойствия.
Споры бактерий разнообразны по форме. Они могут быть круглыми, овальными, с ребрами жесткости. При рассматривании под микроскопом видно, что споры, как и обезвоженный белок, имеют высокий коэффициент преломления света.
Структура зрелой споры состоит из таких слоев:
- Центральный (спороплазма). В состав центрального слоя спор входит цитоплазма, системы белкового синтеза, хромосомы.
- ЦПМ, состоящая из двух слоев. Пространство между двумя слоями спор заполнено гликопептидными полимерами. Они высокочувствительны к лизоциму.
- Оболочка споры состоит в основном из белка. Именно белки обеспечивают высокую устойчивость спор к неблагоприятным условиям. Благодаря им споры способны пролежать сотни лет в ожидании необходимых условий для прорастания.
Отличия вирусов от бактерий
Изучение необходимо для их идентификации, культивирования новых форм, способов получения энергии, изучения взаимодействия микроорганизма со средой, которая его окружает.
Структура клеточных стенок растений
Клеточные стенки растений обладают необычайной прочностью, и в процессе роста растения меняют свою структуру и состав. Основными компонентами клеточных стенок растений являются полисахариды, причем среди них преобладает целлюлоза, которая в значительной мере определяет архитектуру стенки.
Целлюлоза . Этот гомополисахарид является самым распространенным на Земле углеводом (растения образуют в год до 1011 т целлюлозы). Мономерами целлюлозы служат остатки глюкозы, соединенные в длинные цепочки (до 10 000 остатков глюкозы в каждой) с помощью b(1→4)-гликозидных связей (рис. 5.3). В такой молекуле отсутствует полная свобода вращения вокруг 1 С-О- и О- 4С-связей, и полимер приобретает конформацию, благоприятную для образования межцепочечных водородных связей, в случае, когда цепочки располагаются антипараллельно. В результате молекулы целлюлозы объединяются в микрофибриллы толщиной примерно от 10 до 25 нм. Микрофибриллы перевиваются и образуют тонкие нити, которые, в свою очередь, могут обматываться одна вокруг другой, как пряди в канате, формируя макрофибриллы. Каждая макрофибрилла имеет толщину около 0,5 мкм и может достигать в длину 6-8 мкм. Прочность макрофибрилл сопоставима с прочностью равной по толщине стальной проволоки. Кроме того, отдельные участки микрофибрилл имеют упорядоченное строение и придают клеточной стенке кристаллические свойства. Таким образом, можно отметить сложность и высокую упорядоченность целлюлозы в составе клеточных стенок, что неслучайно: этот полимер выполняет защитную и опорную функции в растении.
В таком виде полисахариды недоступны действию собственных ферментов, и целлюлоза не может использоваться растением в качестве резервного вещества. Лишь немногие организмы (некоторые бактерии, грибы, простейшие и редкие животные) обладают ферментными системами, способными расщеплять целлюлозу.
Микро- и макрофибриллы целлюлозы в клеточной стенке растений погружены в матрикс , который также состоит в основном из полисахаридов и меняет свою структуру в процессе роста растения. На начальных этапах развития матрикс состоит из пектиновых веществ , а в дальнейшем в нем появляются ксиланы и различные нейтральные полисахариды («гемицеллюлоза»). Пектиновые вещества представляют собой полимеры a-галактуроновой кислоты, в которых некоторые водородные атомы замещены метильными группами (-СН 3) (рис. 5.3). Ксиланы представляют собой полимеры ксилозы (рис. 5.3).
На более поздних этапах развития, когда происходит одревеснение клеточных стенок, в клетках откладывается лигнин - химически устойчивый полимер, содержащий большое число ароматических колец. Кроме этого, в составе клеточных стенок растений обнаруживаются небольшие количества гликопротеинов, нерастворимых липидных полимеров различного строения и восков.
Клеточные стенки некоторых растений содержат редкие полисахариды, имеющие необычное строение. Например, в стенках и межклеточном
веществе морских красных водорослей содержится сложный гетерополисахарид агар, представляющий собой смесь сульфатированных полисахаридов -агарозы и агаропектина. Агароза построена из чередующихся остатков D-галактозы и 3,6-ангидро-L-лактозы, связанных попеременно b(1→4)- и a(1→3)-связями. Агаропектин имеет более сложное строение: в его состав входят D-галактоза, 3,6-ангидрогалактоза, уроновые кислоты и сульфат. Агар используется в качестве наиболее распространенного уплотнителя для твердых сред, незаменимых в микробиологии, а также в пищевой промышленности для желирования продуктов. Следует отметить, что подавляющее большинство микроорганизмов не способно расщеплять агар, и это одно из главных его преимуществ перед другим уплотнителем питательных сред - желатиной. Агароза находит широкое применение в биохимических исследованиях: она в водной среде образует гель с большими порами, размер которых определяется ее концентрацией. Агарозные гели используются для фрак-ционирования белков и нуклеиновых кислот, а также для иммобилизации клеток.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений
и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию)
белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации
70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные
РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции).
16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав
большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики,
позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов,
бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными
веществами для питания и энергетических потребностей бактерий. Волютин обладает
сродством к основным красителям и легко выявляется с помощью специальных методов
окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное
расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно
прокрашивающихся полюсов клетки.
Нуклеоид
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий
в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка.
Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и
основных белков (гистонов). Обычно в бактериальной клетке содержится одна
хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке
имеются внехромосомные факторы наследственности - плазмиды, представляющие
собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной
стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима
в мазках-отпечатках из патологического материала. В чистых культурах бактерий
капсула образуется реже. Она выявляется при специальных методах окраски мазка
(например, по Бурри-Гинсу), создающих негативное контрастирование веществ
капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов
(экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы
она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует
фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее
увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее
0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать
слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима
в воде.
Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам),
их еще называют гликокаликсом. Кроме синтеза
экзополисахаридов бактериями, существует и другой механизм их образования:
путем действия внеклеточных ферментов бактерий на дисахариды. В результате
этого образуются декстраны и леваны.
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют
собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую
длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят
из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень
со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков
- у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической
мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором,
вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик);
является Н-антигеном. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у
холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии
(перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков
на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков
на противоположных концах клетки.
Пили
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом строения клеточной стенки. Споры образуются при
неблагоприятных условиях существования бактерий (высушивание, дефицит питательных
веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора).
Образование спор способствует сохранению вида и не является способом размножения,
как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие
диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки,
называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium
- веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески
или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с включениями и наследственного аппарата - аналога ядра, называемого нуклеоидом (рис. 2.2). Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры.
Рис. 2.2. Структура бактериальной клетки: 1 - капсула; 2 - клеточная стенка; 3 - цитоплазматическая мембрана; 4 - мезосомы; 5 - нуклеоид; 6 - плазмида; 7 - рибосомы; 8 - включения; 9 - жгутик; 10 - пили (ворсинки)
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной сдерживающая высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов, имеет рецепторы для бактериофагов, бактериоцинов и различных веществ. Наиболее толстая клеточная стенка у грамположительных бактерий (рис. 2.3). Так, если толщина клеточной стенки грамотрицательных бактерий около 15-20 нм, то у грамположительных она может достигать 50 нм и более.
Основу клеточной стенки бактерий составляет пептидогликан. Пептидогликан является полимером. Он представлен параллельными полисахаридными гликановыми цепями, состоящими из повторяющихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью. Эту связь разрывает лизоцим, являющийся ацетилмурамидазой.
К N-ацетилмурамовой кислоте ковалентными связями присоединен тетрапептид. Тетрапептид состоит из L-аланина, который связан с N-ацетилмурамовой кислотой; D-глутамина, который у грамположительных бактерий соединен с L-лизином, а у грамотри-
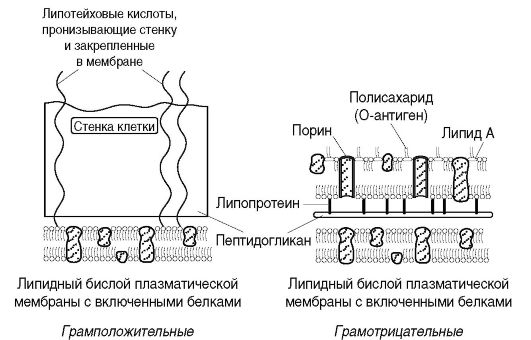 Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
цательных бактерий - с диаминопимелиновой кислотой (ДАП), которая представляет собой предшественник лизина в процессе бактериального биосинтеза аминокислот и является уникальным соединением, присутствующим только у бактерий; 4-й аминокислотой является D-аланин (рис. 2.4).
В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов и белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% массы клеточной стенки. Тетрапептиды разных слоев пептидогликана у грамположительных бактерий соединены друг с другом полипептидными цепочками из 5 остатков глицина (пентаглицина), что придает пептидогликану жесткую геометрическую структуру (рис. 2.4, б). С пептидогликаном ктеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. tekhos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного, с поперечными пептидными сшивками пептидогликана.
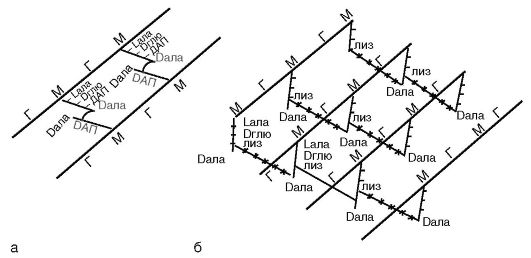 Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого последующая обработка мазка бактерий спиртом вызывает сужение пор в пептидогликане и тем самым задерживает краситель в клеточной стенке.
Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5-10% массы клеточной стенки); они обесцвечиваются спиртом, и при обработке фуксином или сафранином приобретают красный цвет. Это связано с особенностями строения клеточной стенки. Пептидогликан в клеточной стенке грамотрицательных бактерий представлен 1-2 слоями. Тетрапептиды слоев соединены между собой прямой пептидной связью между аминогруппой ДАП одного тетрапептида и карбоксильной группой D-аланина тетрапептида другого слоя (рис. 2.4, а). Кнаружи от пептидогликана расположен слой липопротеина, соединенный с пептидогликаном через ДАП. За ним следуетнаружная мембрана клеточной стенки.
Наружная мембрана является мозаичной структурой, представленной липополисахаридами (ЛПС), фосфолипидами и белками. Внутренний слой ее представлен фосфолипидами, а в наружном слое расположен ЛПС (рис. 2.5). Таким образом, наружная мем-
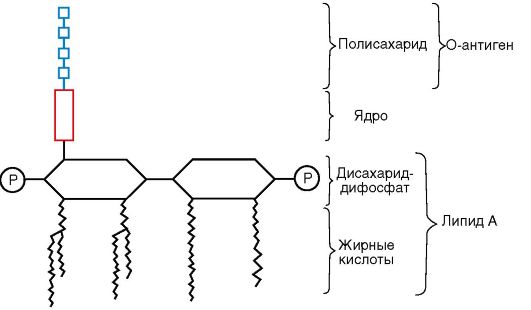 Рис. 2.5.
Структура липополисахарида
Рис. 2.5.
Структура липополисахарида
брана асимметрична. ЛПС наружной мембраны состоит из трех фрагментов:
Липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий. Липид А состоит из фосфорилированных глюкозоаминовых дисахаридных единиц, к которым прикреплены длинные цепочки жирных кислот (см. рис. 2.5);
Ядра, или стержневой, коровой части (от лат. core - ядро), относительно консервативной олигосахаридной структуры;
Высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями.
ЛПС заякорен в наружной мембране липидом А, обусловливающим токсичность ЛПС и отождествляемым поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина, что может вызвать у больного эндотоксический шок. От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота. О-специфическая полисахаридная цепь, отходящая от стержневой части молекулы ЛПС,
состоящая из повторяющихся олигосахаридных единиц, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС связаны представления об О-антигене, по которому можно дифференцировать бактерии. Генетические изменения могут привести к дефектам, укорочению ЛПС бактерий и появлению в результате этого шероховатых колоний R-форм, теряющих О-антигенную специфичность.
Не все грамотрицательные бактерии имеют полноценную О-специфическую полисахаридную цепь, состоящую из повторяющихся олигосахаридных единиц. В частности, бактерии рода Neisseria имеют короткий гликолипид, который называется липоолигосахаридом (ЛОС). Он сравним с R-формой, потерявшей О-антигенную специфичность, наблюдаемой у мутантных шероховатых штаммов E. coli. Структура ЛОС напоминает структуру гликосфинголипида цитоплазматической мембраны человека, поэтому ЛОС мимикрирует микроб, позволяя ему избегать иммунного ответа хозяина.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемыепоринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы с относительной массой до 700 Д.
Между наружной и цитоплазматической мембраной находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, β-лактамазы), а также компоненты транспортных систем.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сфероили протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия Института им. Д. Листера, где они впер-
вые были изучены). L-формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, возвращаясь в исходную бактериальную клетку. L-формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм каждый разделены светлым - промежуточным). По структуре она похожа на плазмолемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов, с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. В отличие от эукариотических клеток, в цитоплазматической мембране бактериальной клетки отсутствуют стеролы (за исключением микоплазм).
Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она окружает наружную часть цитоплазмы бактерий и участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы - АТФазы и др.). При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии. Цитоплазма занимает основной объем бактери-
альной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков.
Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы: 50S и 30S. рРНК - консервативные элементы бактерий («молекулярные часы» эволюции). 16S-рРНК входит в состав малой субъединицы рибосом, а 23S-рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, β-оксимасляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей.
Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в краснофиолетовый цвет, а цитоплазма бактерии - в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронноплотных гранул размером 0,1-1 мкм.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, плотно уложенной наподобие клубка. Нуклеоид бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). У большинства бактерий содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Но у некоторых бактерий имеются две хромосомы кольцевой формы (V. cholerae) и линейные хромосомы (см. раздел 5.1.1). Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК
методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Кроме нуклеоида, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды (см. раздел 5.1.2), представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь. Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка по Бурри- Гинсу, создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, включает большое количество воды. Она препятствует фагоцитозу бактерий. Капсула антигенна: антитела к капсуле вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии.
От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде.
Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам); их еще называют гликокаликсом.
Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие на-
чало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из трех частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (одна пара дисков у грамположительных и две пары у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер.
Жгутики состоят из белка - флагеллина (flagellum - жгутик), являющегося антигеном - так называемый Н-антиген. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий разных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) - нитевидные образования, более тонкие и короткие (3-10 нм * 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Известно несколько типов пилей. Пили общего типа отвечают за прикрепления к субстрату, питание и водно-солевой обмен. Они многочисленны - несколько сотен на клетку. Половые пили (1-3 на клетку) создают контакт между клетками, осуществляя между ними передачу генетической информации путем конъюгации (см. главу 5). Особый интерес представляют пили IV типа, у которых концы обладают гидрофобностью, в результате чего они закручиваются, эти пили называют еще кудряшками. Располага-
ются они по полюсам клетки. Эти пили встречаются у патогенных бактерий. Они обладают антигенными свойствами, осуществляют контакт бактерии с клеткой-хозяином, участвуют в образовании биопленки (см. главу 3). Многие пили являются рецепторами для бактериофагов.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки. Спорообразующие бактерии рода Bacillus, у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие бактерии, у которых размер споры превышает диаметр клетки, отчего они принимают форму веретена, называются клостридиями, например бактерии рода Clostridium (от лат.Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нельсена в красный, а вегетативная клетка - в синий цвет.
Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор бывает овальной и шаровидной, расположение в клетке - терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены) и центральное (у сибиреязвенной бациллы).
Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома бактериальной вегетативной клетки отделяются, окружаясь врастающей цитоплазматической мембраной, - образуется проспора.
В протопласте проспоры находятся нуклеоид, белоксинтезирующая система и система получения энергии, основанная на гликолизе. Цитохромы отсутствуют даже у аэробов. Не содержится АТФ, энергия для прорастания сохраняется в форме 3-глицеринфосфата.
Проспору окружают две цитоплазматические мембраны. Слой, окружающий внутреннюю мембрану споры, называется стенкой споры, он состоит из пептидогликана и является главным источником клеточной стенки при прорастании споры.
Между наружной мембраной и стенкой споры формируется толстый слой, состоящий из пептидогликана, имеющего много сшивок, - кортекс.
Кнаружи от внешней цитоплазматической мембраны расположена оболочка споры, состоящая из кератиноподобных белков, со-
держащих множественные внутримолекулярные дисульфидные связи. Эта оболочка обеспечивает резистентность к химическим агентам. Споры некоторых бактерий имеют дополнительный покров -экзоспориум липопротеиновой природы. Таким образом формируется многослойная плохо проницаемая оболочка.
Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция.
Спора долго может сохраняться из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет.
В благоприятных условиях споры прорастают, проходя три последовательные стадии: активации, инициации, вырастания. При этом из одной споры образуется одна бактерия. Активация - это готовность к прорастанию. При температуре 60-80 °С спора активируется для прорастания. Инициация прорастания длится несколько минут. Стадия вырастания характеризуется быстрым ростом, сопровождающимся разрушением оболочки и выходом проростка.
1)СТРОЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ:
Клетка прокариотических организмов имеет сложное строго упорядоченное строение и обладает принципиальными особенностями ультраструктурной организации и химического состава.
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и яд¬ра, называемого нуклеоидом. Имеются дополни¬тельные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях спо¬собны образовывать споры.
Функции клеточной стенки:
1. Определяет и сохраняет постоянную форму клетки.
2. Защищает клетку от механических и осмотичес¬ких сил внешней среды.
3. Участвует в регуляции роста и деления клеток.
4. Обеспечивает коммуникацию с внешней средой через каналы и поры.
5. Несет на себе специфические рецепторы для бактериофагов.
6. Определяет во многом антигенную характеристику бактерий (природу и спе¬цифичность О- и К-антигенов).
8. Нарушение синтеза клеточной стенки бактерий является главной причиной их L-трансформации.
Метод выявления клеточной стенки - электронная микроскопия, плазмолиз.Термин «оболочка» в настоящее время используется для обозначения клеточной стенки и капсулы бактерий или только клеточной стенки, цитоплазматическая мембрана не входит в состав оболочки и относится к протопласту.
Основным компонентом клеточной стенки бактерий является пептидогликан, или муреин (лат. murus - стенка), - опорный полимер, имеющий сетчатую структуру и образующий ригидный (жесткий) наружный каркас бактериальной клетки.
2)СПОРА БАКТЕРИЙ, СТРОЕНИЕ, НАЗНАЧЕНИЕ, ОТЛИЧИЕ ОТ СПОР ГРИБОВ.
Споры бактерий - тельца круглой или овальной формы, которые образуются внутри некоторых бактерий в определенные стадии их существования или при ухудшении условий окружающей среды. Размер, форма и расположение спор в клетке - признак относительно постоянный, характерный для некоторых видов бактерий. Споры бактерий устойчивы к различным физическим и химическим воздействиям, не окрашиваются обычными анилиновыми красителями и сохраняются в течение нескольких лет, не утрачивая свойства прорастать в вегетативную форму, что имеет значение в эпидемиологии ряда заболеваний. Споры бактерий погибают после прогревания в автоклаве при t° 120° в течение 30 минут или при обработке сухим жаром при t° 160- 180° в течение часа.
В зрелой споре различимы: центральный, плохо окрашиваемый участок (спороплазма), двухслойная ЦПМ и оболочка споры.
Спороплазма (протопласт споры) включает цитоплазму, бактериальную хромосому, системы белкового синтеза и некоторые другие (например, анаэробного энергообразования).
Оболочка споры двухслойная: пространство между слоями заполняют гликопептидные полимеры, сходные с пептидогликанами, образующие сетчатую структуру (кортекс), проявляющую высокую чувствительность к лизоциму.
Внутренний слой (стенка споры) образован пептидогликанами, аналогичными таковым \ вегетирующей клетки.
Внешний слой (собственно оболочка) образуют кератиноподобные белковые структуры с низкой проницаемостью.
Экзоспориум. У некоторых бактерий материнская клетка образует экзоспориум - двух-трёхслойное желатинообразное покрытие образованное липопротеинами и углеводами и во многом аналогичное капсуле бактерий. При созревании споры экзоспориум может сохраняться в виде пустого и отстающего от споры «мешка»
Споры представляют собой своеобразные покоящиеся клетки; у них чрезвычайно низкая метаболическая активность, но они обладают высокой устойчивостью к высушиванию, действию повышенной температуры и различных химических веществ. Высокую резистентность спор к действию указанных факторов связывают с присутствием в оболочке большого количества кальциевой соли дипиколиновой кислоты. Споры сильно преломляют свет, поэтому они хорошо заметны в неокрашенных препаратах.
Споры грибов,в отличие от бактерий,служат для размножения. У грибов споры могут развиваться внутри специальных споровместилищ (эндогенно) или на конце особых выростов мицелия - конидиеносцах (экзогенно).
3)ОСНОВНЫЕ ЭЛЕМЕНТЫ БЕЛОКСИНТЕЗИРУЮЩЕЙ СИСТЕМЫ БАКТЕРИЙ.
Синтез белка осуществляется с помощью сложной белоксинтезирующей системы. В ее состав входят следующие компоненты:
1. Рибосомные субъединицы 30S и 50S, которые у прокариот и в митохондриях и хлоропластах эукариот образуют рибосому 70S; или субъединицы 40S и 60S, образующие у эукариот рибосому 80S.
2. Матричная РНК (мРНК).
3. Полный комплект двадцати аминоацил-тРНК, для образования которых необходимы соответствующие аминокислоты, аминоацил-тРНК-синтетазы, тРНК и АТФ. Аминоацил-тРНК (аа тРНК) - это заряженная энергией и связанная с тРНК аминокислота, готовая для подвоза к рибосоме и включения в синтезирующийся на ней полипептид.
4. Белковые факторы инициации (у прокариот - IF-1, IF-2, IF-3).
5. Белковые факторы элонгации (у прокариот - EF-Tu, EF-Ts, EF-G).
6. Белковые факторы терминации (у прокариот - RF-1, RF-2, RF-3).
7. Некоторые другие белковые факторы (ассоциации, диссоциации субъединиц, высвобождения и пр.).
8. Гуанозинтрифосфат (ГТФ).
9. Неорганические катионы в определенной концентрации.
4)ПРИНЦИП ФАЗОВО-КОНТРАСТНОЙ МИКРОСКОПИИ
При микроскопии неокрашенных микроорганизмов, отличающихся от окружающей среды только по показателю преломления, изменения интенсивности света (амплитуды) не происходит, а изменяется только фаза прошедших световых волн. Поэтому глаз этих изменений заметить не может и наблюдаемые объекты выглядят малоконтрастными, прозрачными. Для наблюдения таких объектов используют фазово-контрастную микроскопию, основанную на превращении невидимых фазовых изменений, вносимых объектом, в амплитудные, различимые глазом.
Фазово-контрастное устройство может быть установлено на любом световом микроскопе и состоит из:
1) набора объективов со специальными фазовым пластинками;
2) конденсора с поворачивающимся диском. В нем установлены кольцевые диафрагмы, соответствующие фазовым пластинкам в каждом из объективов;
3) вспомогательного микроскопа.
Благодаря применению этого способа микроскопии контраст живых неокрашенных микроорганизмов резко увеличивается и они выглядят темными на светлом фоне (позитивный фазовый контраст) или светлыми на темном фоне (негативный фазовый контраст).
5)ПРИНЦИП ТЕМНОПОЛЬНОЙ МИКРОСКОПИИ
Темнопольная микроскопия позволяет наблюдать живые бактерии. Для темнопольной микроскопии используют темнопольный конденсор, выделяющий контрастирующие структуры неокрашенного материала
Темнопольная микроскопия. Микроскопия в темном поле зрения основана на явлении дифракции света при сильном боковом освещении взвешенных в жидкости мельчайших частиц (эффект Тиндаля). Эффект достигается с помощью параболоид-или кардиоидконденсора, которые заменяют обычный конденсор в биологическом микроскопе.(препарат освещается сбоку косыми пучками лучей.не попадающими в объектив.)
6)ОСНОВНЫЕ ОТЛИЧИЯ МОЖДУ ПРОКАРИОТАМИ И ЭУКАРИАОТАМИ.
У прокариот нет ядра, кольцевая ДНК (кольцевая хромосома) расположена прямо в цитоплазме (этот участок цитоплазмы называется нуклеоид).
У эукариот есть оформленное ядро (наследственная информация [ДНК] отделена от цитоплазмы ядерной оболочкой).
1) Раз у прокариот нет ядра, то нет и митоза/мейоза. Бактерии размножаются делением надвое.
2) У прокариот из органоидов имеются только рибосомы (мелкие, 70S), а у эукариот кроме рибосом (крупных, 80S) имеется множество других органоидов: митохондрии, эндоплазматическая сеть, клеточный центр, и т.д.
3) Клетка прокариот гораздо меньше клетки эукариот: по диаметру в 10 раз, по объему – в 1000 раз.
Прокариоты отличаются от эукариот по ряду основных признаков .
1.Отсутствие истинного дифференцированного ядра (ядерной мембраны).
2.Отсутствие развитой эндоплазматической сети, аппарата Гольджи.
3.Отсутствие митохондрий, хлоропластов, лизосом.
4.Неспособность к эндоцитозу (захвату частиц пищи).
5.Клеточное деление не связано с циклическими изменениями строения клетки.
6. Значительно меньшие размеры (как правило). Большая часть бактерий имеет размеры 0,5- 0,8 микрометров (мкм ) х 2- 3 мкм.
7)ПРИЗНАКИ ПРИМЕНЯЕМЫЕ ДЛЯ ИДЕНТИФИКАЦИИ МИКРООРГАНИЗМОВ.
Морфологические-форма,величина,характер расположения микробов друг к другу.
Тинкториальные –способность микробов окрашиваться различными красителями.
Культуральные-особенности роста микробов на жидких и плотных питательных средах.
Биохимические-способность организмов потреблять различные вещества (углеводы,белки и др)с образованием конечных продуктов или наличие у микроорганизмов различных ферментов(уретаза,цистиназа и др)
Антигенные-особенности хим.состава микробных клеток (антигенной структуры) выявляемые с помощью спец диагностич.сывороток чувствительность к спец. фагам.
8)ОПРЕДЕЛЕНИЯ ВСЕХ МЕТОДОВ МИКРОБИОЛОГИЧЕСКОЙ ДИАГНОСТИКИ
Микроскопический. С помощью микроскопии нативного патологического материала определяют вид возбудителя заболевания по форме, взаиморасположению и способности окрашиваться определенными кра¬сителями.
Бактериологический. Этот метод основан на выделении чистой культуры возбудителя заболевания и его идентификации (определение вида микроба).
Серологический. Метод основан на определении специфических антител в крови больных или переболевших инфекционными заболева¬ниями к соответствующим возбудителям с помощью различных реакций (агглютинации, пассивной гемагглютинации, связывания комплемента, преципитации и других).
Биологический. В основе этого метода лежит заражение лабораторных животных исследуемым материалом от больного с целью воспроизведения у них инфекционного заболевания или последующего выделения возбудителя.
Аллергический. С помощью этого метода обнаруживают повышен¬ную чувствительность макроорганизма к определенным возбудителям инфекционных заболеваний или продуктам их жизнедеятельности. Для аллергических проб используют препараты, которые называются аллергенами.
9)СТРУКТУРА ЖГУТИКА БАКТЕРИЙ
Жгутик - спирально изогнутая полая нить, образованная субъединицами флагеллина. У разных бактерий толщина жгутиков варьирует от 12 до 18 нм, что составляет не более 1/10 диаметра жгутиков водорослей и простейших. Жгутики также различают по длине и диаметру витка. Место прикрепления жгутика к бактериальной клетке имеет сложное строение и состоит из базальной структуры и так называемого «крюка» (рис. 4-2). У грамположительных бактерий в состав базальной структуры входит одна пара, а у грамотрицательных бактерий - две пары колец. Кольца играют роль «приводного диска» и «подшипника». Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор). У спирохет за движение ответственна особая органел-ла - осевая нить, состоящая из двух рядов бактериальных жгутиков, расположенных продольно внутри клетки.
Жгутики бактерий состоят из трёх субструктур:
Филамент (фибрилла, пропеллер) - полая белковая нить толщиной 10-20 нм и длиной 3-15 мкм, состоящая из флагеллина, субъединицы которого уложены по спирали. Полость внутри используется при синтезе жгутика - он происходит в направлении от плазматической мембраны. По полости к собираемому в настоящий момент участку переносятся субъединицы флагеллина.
Крюк - более толстое, чем филамент (20-45 нм), белковое (не флагеллиновое) образование.
Базальное тело (трансмембранный мотор)
10)КЛАССИФИКАЦИЯ БАКТЕРИЙ ПО КОЛИЧЕСТВУ И ВЗАИМРАСПОЛОЖЕНИЮ ЖГУТИКОВ
Расположение жгутиков - характерный признак, имеющий таксономическое значение. У некоторых бактерий жгутики расположены по всей поверхности клеточной стенки (например, у бактерий рода Proteus), такие бактерии известны как перитрихи [от греч. peri-, вокруг, + trichos, волос].
Некоторые бактерии снабжены только одним толстым жгутиком (например, представители рода Vibrio), они известны как монотрихи.
Политрихи - бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии.
Монополярно-политрихиальное расположение жгутиков имеют лофотрихи [от греч. lophos, пучок, + trichos, волос], к ним, например, относят представителей рода Pseudomonas.
Биполярно-политрихиальное жгутикование имеют амфитрихи [от греч. amphi-, двусторонний, + trichos, волос] (например, бактерии рода Spirillum).
11)МЕТОДЫ ОПРЕДЕЛЕНИЯ ПОДВИЖНОСТИ БАКТЕРИЙ
Подвижность бактерий определяют микроскопией препаратов в «раздавленной» или «висячей» капле. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные - по уколу) или посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют её), либо определяют способность бактерий давать «феномен роения».
12)СТРУКТУРА ПЕПТИДОГЛИКАНА
(Муреин) Пептидогликан образует упорядоченную структуру ячеистого строения, построенную из N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных β-1,4-гликозидными связями. Остатки N-ацетилмурамовой кислоты сшиты между собой при помощи коротких пептидов (сшивка производится ферментом транспептидазой). Типично пептидная цепочка содержит L-аланин, D-глутаминовую кислоту, мезо-диаминопимелиновую кислоту, L-лизин, D-аланин (следует отметить, что мезо-диаминопимелиновая кислота и D-аминокислоты в составе клеточных структур встречаются только в прокариотических клетках). Такая трёхмерная структура сообщает клеточной стенке бактерий прочность и защиту от осмотического лизиса.
13) ПРИНЦИП ОКРАСКИ БАКТЕРИЙПО ГРАМУ.ОТЛИЧИЕ КЛЕТОЧНОЙ СТЕНКИ ГРАМОПОЛОЖИТЕЛЬНЫХ БАКТЕРИЙОТ ГРАМОТРИЦАТЕЛЬНЫХ.
Принцип окраски по Граму основан на том, что положительно красящиеся по Граму микробы имеют в своем поверхностном слое вещества (рибонуклеат магния с белками), которые при окрашивании парарозанилиновыми красками (генцианвиолет, кристаллвиолет, метилвиолет, викторияблау) в присутствии иода дают прочные соединения, не разрушающиеся спиртом. Микробы, отрицательно красящиеся по Граму, в своем составе не имеют таких веществ и потому при окрашивании этими красками в присутствии иода не дают прочных соединений, и при действии спирта обесцвечиваются. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
14)В КАКИХ СЛУЧАЯХ ИМСПОЛЬЗУЮТСЯ ТЕМИНЫ: «БАКТЕРИИ», «БАЦИЛЛЫ», «КЛОСТРИДИИ»
Термин «Бактерии» используется для прокариотических микрооганизмов, чаще всего одноклеточных.
Термин «Бациллы» используется для грамположительных палочковидных бактерий,образующих внутриклеточные споры.
Темин «клостридии» используется для обозначения рода грамположительных облигатно-анаэробных бактерий способных продуцировать эндоспоры.
15)ЧТО ТАКОЕ LФОРМЫ БАКТЕРИЙ,В КАКИХ СЛУЧАЯХ ОНИ ВОЗНИКАЮТ
L-формы - бактерии, частично или полностью лишённые клеточной стенки, но сохранившие способность к развитию. Было показано, что L-формы возникают спонтанно или индуцировано - под воздействием агентов, блокирующих синтез клеточной стенки: антибиотиков (пенициллины циклосерин, цефалоспорины, ванкомицин), ферментов (лизоцим, амидаза, эндопептидаза) ультрафиолетовых и рентгеновских лучей, аминокислоты глицина.
L-формы образуются в результате несбалансированного роста нормальных бактериальных клеток в длину и в толщину и поэтому полиморфны. В культурах L-форм обнаруживаются шаровидные, нитевидные или вовсе бесструктурные клетки размером от 0,2 до 50 мкм. Они спокойно проходят через бактериальные фильтры и легко разрушаются при механических воздействиях. В отличие от нормальных клеток L-формы часто содержат крупные вакуоли. Их метаболическая активность очень низкая. Клеточное деление происходит нестандартно, за счёт образования элементарных тел путём отпочкования от поверхности клетки или от мембраны вакуоли.
16)МЕТОДЫ ВЫЯВЛЕНИЯ СПОР У БАКТЕРИЙ
Обнаружение спор.
Благодаря толщине свое оболочки и плотности содержимого, споры остаются неокрашенными при обработке препарата анилиновыми красителями простым методом или сложным по Граму.
При окраске по Граму или по Леффлеру спора внутри окрашенной цитоплазмы микробной клетки выглядит как зернышко круглой или овальной формы, сильно преломляющее свет.
Существует несколько методов окраски спор (по Циль – Нильсену, по Ганзену, по Ожешко и др.) позволяющих достигнуть контрастной окраски спор в цитоплазме.
Методика окраски микропрепарата:
1) На фиксированный препарат накладывается полоска фильтровальной бумаги (для защиты препарата от оседающих кристаллов красителя) и на нее наливается карболовый фуксин Циля. Препарат осторожно прогревается в течение 3 – 4 минут над пламенем горелки. По мере испарения жидкости краситель добавляется.
2) Фильтровальная бумага снимается и на мазок наносится 2 – 3 капли 5% раствора кислоты (серной, соляной, азотной или уксусной) на 30 секунд.
3) Препарат тщательно промывается струей холодной воды и высушивается.
4) Докрашивается раствором метиленовой сини Леффлера в течение 1 – 2 минут.
5) Препарат промывается струей воды, высушивается и микроскопируется. На голубом фоне цитоплазмы видны сиренево – красные споры.
Этот метод позволяет обнаружить споры не только в процессе их формирования внутри клетки, но и после того как сформированная спора высыпалась из разрушившейся микробной клетки.
Приготовление карболового фуксина Циля:10 мл. насыщенного спиртового раствора фуксина растворяют в 100 мл 5% раствора карболовой кислоты.
17)ЧТО ТАКОЕ НУМЕРИЧЕСКАЯ ТАКСОНОМИЯ
Таксономия нумерическая
метод феносистематики, состоящий в получении максимальной информации об иссл. микробе. Проводится в несколько этапов: 1) отбор иссл. к-р и типовых штаммов; 2) выбор признаков и их кодирование; 3) определение признаков; 4) компьютерный анализ результатов; 5) интерпретация результатов на основе степени сходства и различий. При кодировании все признаки уравниваются или располагаются иерархически.
Нумерическая (численная) таксономия основывается на использовании максимального количества сопоставляемых признаков и математическом учете степени соответствия. Больщое число сравниваемых фенотипических признаков и принцип их равной значимости затрудняло классификацию.
18)ЧТО ТАКОЕ КОНСТИТУТИВНЫЕ,ИНДУЦИБЕЛЬНЫЕ, РЕПРЕСИБЕЛЬНЫЕ ФЕРМЕНТЫ
1. конститутивные - синтез которых происходит с определенным постоянством и не зависит от субстрата и конечного продукта
2. индуцибельные- синтез которых усиливается при наличии в окружающей среде субстрата для этих ферментов
3.репресибельные- синтез которых подавляется при наличии конечного продукта реакции в окружающей среде
19)ОТЛИЧАЕ МЕЖДУ ЯДЕРНЫМ АППАРАТОМ ПРОКАРИОТ И ЭУКАРИОТ
Принципиальное и наиболее заметное различие между этими двумя группами организмов заключается в том, что эукариоты имеют особый органоид – ядро, т. е. генетический материал обособлен и отделен от цитоплазмы ядерной оболочкой. Наследственная информация заключена в хромосомах, содержащих ДНК и особые белки. Деление происходит в результате сложного процесса, называемого митозом. Однако неправильно будет сказать, что прокариоты не имеют ядра, так как у них есть ядроподобные структуры – нуклеоиды. Более корректное выражение: прокариоты не имеют оформленного ядра, так как нукле-оиды имеют более простое строение и не отделены от цитоплазмы мембраной. Наследственную информацию несет одна хромосома, представляющая собой длинную молекулу ДНК.
20)СТРУКТУРА И ФУНКЦИИ ЦИТОПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ
Основу плазмалеммы, как и других мембран в клетках (например, митохондрий, пластид и т. д.), составляет слой липидов, имеющий два ряда молекул (рис. 1). Поскольку молекулы липидов полярны (один полюс у них гидрофилен, т. е. притягивается водой, а другой гидрофобен, т. е. отталкивается от воды), то и располагаются они в определенном порядке. Гидрофильные концы молекул одного слоя направлены в сторону водной среды - в цитоплазму клетки, а другого слоя - наружу от клетки - в сторону межклеточного вещества (у многоклеточных) или водной среды (у одноклеточных).
Выделяют периферические белки (они расположены только по внутренней или наружной поверхности мембраны), интегральные (они прочно встроены в мембрану, погружены в нее, способны менять свое положение в зависимости от состояния клетки). Функции мембранных белков: рецепторная, структурная (поддерживают форму клетки), ферментативная, адгезивная, антигенная, транспортная.
Схема строения элементарной мембраны жидкостно-мозаичная: жиры составляют жидкокристаллический каркас, а белки мозаично встроены в него и могут менять свое положение.
Молекулы белков мозаично встроены в бимолекулярный слой липидов. С внешней стороны животной клетки к липидам и молекулам белков плазмалеммы присоединяются молекулы полисахаридов, образуя гликолипиды и гликопротеины.Эта совокупность формирует слой гликокаликса. С ним связана рецепторная функция плазмалеммы (см. ниже); также в нем могут накапливаться различные вещества, используемые клеткой. Кроме того, гликокаликс усиливает механическую устойчивость плазмалеммы.
1) барьерная (отграничение внутреннего содержимого клетки);
2) структурная (придание определенной формы клеткам в соответствии с выполняемыми функциями);
3) защитная (за счет избирательной проницаемости, рецепции и антигенности мембраны);
4) регуляторная (регуляция избирательной проницаемости для различных веществ (пассивный транспорт без затраты энергии по законам диффузии или осмоса и активный транспорт с затратой энергии путем пиноцитоза, эндо- и экзоцито-за, работы натрий-калиевого насоса, фагоцитоза)).
5) адгезивная функция (все клетки связаны между собой посредством специфических контактов (плотных и неплотных));
6) рецепторная (за счет работы периферических белков мембраны). Существуют неспецифические рецепторы, которые воспринимают несколько раздражителей (например, холодовые и тепловые терморецепторы), и специфические, которые воспринимают только один раздражитель (рецепторы световоспринимающей системы глаза);
7) электрогенная (изменение электрического потенциала поверхности клетки за счет перераспределения ионов калия и натрия (мембранный потенциал нервных клеток составляет 90 мВ));
8) антигенная: связана с гликопротеинами и полисахаридами мембраны. На поверхности каждой клетки имеются белковые молекулы, которые специфичны только для данного вида клеток. С их помощью иммунная системы способна различать свои и чужие клетки. Обмен веществ между клеткой и окружающей средой осуществляется разными способами - пассивными и активными.
21)ФУНКЦИИ КЛЕТОЧНОЙ СТЕНКИ БАКТЕРИЙ
сновные функции клеточной стенки следующие.
Клеточная стенка защищает бактерии от внешних воздействий, придаёт им характерную форму, поддерживает постоянство внутренней среды и участвует в делении.
Через клеточную стенку бактерий осуществляется транспорт питательных веществ и выделение метаболитов,
На поверхности клеточной стенки располагаются рецепторы для бактериофагов, бактериоци-нов и различных химических веществ.
Структура и состав элементов клеточной стенки определяет антигенную характеристику бактерий (по структуре О- и Vi-Аг).
Клеточная стенка способна по-разному воспринимать красители; на этом основаны тинкториальные свойства бактерий. Нарушение синтеза компонентов клеточной стенки приводит к гибели бактерии или образованию 1-форм.
22)ТИПЫ ДЫХАНИЯ БАКТЕРИЙ,СУЩНОСТЬ И РАЗНИЦА
Дыхание (или биологическое окисление) - это сложный процесс, который сопровождается выделением энергии, необходимой микроорганизмам для синтеза различных органических соединений.
Облигатные (строгие) аэробы развиваются при наличии в атмосфере 20% кислорода (микобактерии туберкулеза), содержат ферменты, с помощью которых осуществляется перенос водорода от окисляемого субстрата к кислороду воздуха.
Микроаэрофилы нуждаются в значительно меньшем количестве кислорода, и его высокая концентрация хотя и не убивает бактерии, но задерживает их рост (актиноисцеты, бруцеллы, лептоспиры).
Факультативные анаэробы могут размножаться как в присутствии, так и в отсутствие кислорода (большинство патогенных и сапрофитных микробов - возбудители брюшного тифа, паратифов, кишечная палочка).
Облигатные анаэробы - бактерии, для которых наличие молекулярного кислорода является губительным (клостри-дии столбняка, ботулизма).
Аэробные бактерии в процессе дыхания окисляют различные органические вещества (углеводы, белки, жиры, спирты, органические кислоты и пр.).
Дыхание у анаэробов происходит путем ферментации субстрата с образованием небольшого количества энергии. Процессы разложения органических веществ в безкислородных условиях, сопровождающиеся выделением энергии, называют брожением.
23) СТРОЕНИЕ И ФУНКЦИИ ЛИПОПОЛИСАХАРИДА КЛЕТОЧНОЙ СТЕНКИ
Липополисахарид (ЛПС; англ. lipopolysaccharide) - макромолекула, состоящая из полисахарида, ковалентно соединённого с липидом, основной компонент клеточной стенки грамотрицательных бактерий.
ЛПС включает 3 ковалентно-связанных компонента:
Липид А
Центральный олигосахарид
О -антиген
ЛПС обеспечивает структурную целостность бактериальной клетки и защищает мембрану от агрессивных воздействий окружающей среды. Отрицательный заряд ЛПС повышает общий отрицательный заряд бактерии и стабилизирует её мембрану. Действие многих антибиотиков направлено на повреждение ЛПС.Единственной грамположительной бактерией с липополисахаридом является Listeria monocytogenes.
24)ЧТО ТАКОЕ «ВИД» МИКРООРГАНИЗМА
Вид - совокупность микроорганизмов, имевших общий корень происхождения, сходный генотип (более 80% гомологии ДНК) и максимально близкие фенотипические признаки и свойства (А.И.Коротяев).
25)ЧТО ТАКОЕ АРХЕБАКТЕРИИ
Архебактерии, субцарство царства прокариот (организмов, клетки которых не имеют оформленного ядра). Архебактерии по составу ДНК и РНК и по биохимии значительно отличаются от бактерий. Полагают, что эти организмы напоминают древние бактерии, которые первыми выросли в экстремальном окружении, в богатых серой морских глубинах.
У архебактерий уникальные протеиноподобные стенки клеток и клеточные мембраны и характерные рибосомы.
К архебактериям относятся метанопроизводящие бактерии, которые используют в качестве пищи такие простые органические соединения, как метанол и ацетат, комбинируя их с диоксидом углерода и водородом воздуха и выделяя метан как побочный продукт. Бактерии, которые живут в горячих источниках и в засоленных зонах, пользуются различными способами получения пищи и энергии, они способны "переваривать" минералы вместо органических соединений. Они могут быть аэробы и анаэробы. Некоторые из живущих в горячих источниках терпимы к температуре до 88 С и кислотности до рН 0,9. Один вид Thermoplasta, вероятно, родствен предшественнику ядра и цитоплазмы более продвинутых клеток эукариотов. Некоторые систематики относят архебактерии к организмам, которые отличаются от всего живущего на Земле, и выделяют их в отдельную группу.