Блок 2. ДНК. Вопросы 5,6,7.
Структура ДНК. Модель Дж. Уотсона и Ф. Крика. Свойства и функции наследственного материала.
Самовоспроизведение генетического материала. Репликация ДНК.
Организация наследственного материала у про- и эукариот. Классификация нуклеотидных последовательностей в геноме эукариот (уникальные, среднеповторяющиеся, высокоповторяющиеся).
В 1868 г. швейцарский химик Ф. Мишер обнаружил в клеточных ядрах, изолированных из гноя, а позже из спермиев лосося вещество, которое он назвал «нуклеином» (от лат. nucleus - ядро). Впоследствии Р. Альтманн (1889 г.) сообщил, что выделенный Ф. Мишером «нуклеин» состоит из двух фракций - белковой и нуклеиновых кислот. Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразумевается их нуклеотидная последовательность) и трехмерной структурой. Интерес к структуре ДНК усилился, когда в начале XX в. возникло предположение, что ДНК , возможно, представляет собой генетический материал. В 1952 году Чаргаффом было открыто правило комплиментарности, назнаванное впоследствии именем создателя. Оно заключается в том, что:
- Количество аденина равно количеству тимина, а гуанина - цитозину: А=Т, Г=Ц.
- Количество пуринов равно количеству пиримидинов: А+Г=Т+Ц.
- Количество оснований с аминогруппами в положении 6 равно количеству оснований с кетогруппами в положении 6: А+Ц=Г+Т.
Впоследствии, Уилкинсоном была получена рентгенограмма ДНК. А еще несколько позже Уотсон и Крик в 1953 году предложили собственную модель ДНК, за которую вместе с Уилкинсоном были удостоены Нобелевской премии в 1962 году.
Основные принципы строения ДНК.
1. Мономер ДНК-нуклеотид, состоящий из азотистого основания, дезоксирибозы и остатка фосфорной кислоты. Азотистые основания могут быть пуриновые А,Г или пиримидиновые Ц,Т.
2. Азотистые основания присоединяются к С1 атому углерода в молекуле пентозы, а к С5 присоединяется фосфат. У третьего атома всегда есть группа ОН.
3. При взаимодействии фосфата одного нуклеотида с гидроксилом дезоксирибозы другого устанавливается фосфодиэфирная связь.
4.Соединение нуклеотидов происходит через ОН пентозы в положение С3 и фосфата последующего нуклеотида.
5. ДНК есть двойная полинуклеотидная цепь. Две полинуклеотидных цепи связаны между собой водородными связями по принципу комплиментарности, А-Т и Г-Ц. Между А и Т две водородные связи, между Т и Ц три водородные связи.
6. Антипараллельность. 5 конец одной цепи соединяется с 3 концом другой цепи.
7.Диамет спирали ДНК равен 2 нм, а длина шага – 3,4 нм. На каждый виток приходится 10 пар нуклеотидов.
8. Первичная структура - полинуклеотидная цепь.
Вторичная структура - две комплиментарные друг другу антипараллельные полинуклеотидные цепи.
Третичная структура - трехмерная спираль.
9. ДНК обладает способностью к репликации.
РЕПЛИКАЦИЯ.

1 - матричные цепи ДНК; 2 - фермент геликаза, разделяющий цепи матричной ДНК; 3 - ДСБ-белки, препятствующие воссоединению цепей ДНК; 4 - праймаза; 5 - РНК-затравка (синтезируется РНК-полимеразой - праймазой); 6 - ДНКполимераза, синтезирующая дочерние цепи; 7 - лидирующая дочерняя цепь ДНК; 8 - лигаза, соединяющая фрагменты Оказаки отстающей цепи ДНК; 9 - фрагмент Оказаки (150-200 нуклеотидов); 10 – топоизомераза
Синтез новой молекулы ДНК осуществляется полуконсервативным способом. Это значит, что дочерняя молекула будет содержать одну материнскую и одну вновь синтезированную цепь. Так как синтез ДНК происходит на одноцепочечной матрице, ему предшествует обязательное временное разделение двух цепей, с образованием репликативной вилки. С помощью электронного микроскопа выявили, что область репликации имеет вид глазка внутри нереплицированной ДНК (репликационный глазок, состоящий примерно из 300 нуклеотидов).
Репликон – фрагмент ДНК от точки начала репликации до точки ее окончания.
Чтобы расплести спираль ДНК нужны особые ферменты(белки). В репликации принимают участие несколько ферментов, каждый из которых выполняет свою функцию.
ДНК-хеликазы(геликазы) разрывают водородные связи между основаниями, разделяют цепи и продвигают репликативную вилку.
Дестабилизирующие белки удерживают цепи.
ДНК –топоизомераза. Вспомним, что ДНК представляет собой спираль. Соответственно, чтобы вилка могла двигаться вперед, спираль должна быстро раскручиваться. Но это потребует большой потери энергии. На самом деле, этого все же не происходит. Этому и способствуют ДНК-топоизомеразы. Они вносят в цепьодно- и двухцепочечные разрывы, позволяющие цепям разделиться, а затем ликвидируют эти разрывы. Благодаряу одна из цепей ДНК начинает вращаться вокруг второй цепи. Также они участвуют в расцеплении колец, образующихся при репликации кольцевых ДНК.
Синтез цепей ДНК происходит с помощью ДНК-полимеразы. Но у этого фермента есть особенность. Он способен добавлять нуклеотиды к 3 концу уже имеющейся цепи. Такую заранее образованную цепь называют затравкой, которую синтезирует праймаза. РНК-затравка отличается от остальной цепи ДНК, так как в ней есть рибоза. Размер затравки невелик. Выполнившая свою функцию затравка удаляется специальным ферментом, а образованная при этом брешь ликвидируется ДНК-полимеразой (в данном случае вместо затравки она использует 3ОН конецсоседнего фрагмента ДНК).
Репликация ДНК предполагает, что синтез двух цепей происходит одновременно. Но на самом деле все происходит не совсем так. Вспомним, что цепи антипараллельны. А синтез новой цепи может происходить только в направлении от 5 конца к 3. Поэтому непрерывно синтез происходит только на одной цепи (лидирующей). На второй (отстающей) он происходит фрагментами Оказаки. Синтез каждого из фрагментов осуществляется с помощью РНК-затравки. Затем затравки удаляются, бреши заполняются ДНК-полимеразой и фрагменты сшиваются ферментом лигазой .
Структурно-функциональная организация ДНК у про- и эукариот
Изучите таблицы, перепишите их в рабочую тетрадь.
| Признаки | Прокариоты | Эукариоты |
| Количество генов | 4 тыс. (Е. coli) | Около 30 тыс. (человек) |
| Количество ДНК | 4 млн пар нуклеотидов | 3-7 млрд пар нуклеотидов |
| Кодирующие последовательности ДНК | Более 90% | Менее 10% |
| Связь ДНК с гистонами | Отсутствует | Формирует нуклеосомы |
| Укладка ДНК | Кольцевая, содержит 100 петель по 40 тыс. пар нуклеотидов | Линейная с замкнутыми в теломеры концами, имеет 4 уровня спирализации |
| Количество репликонов | Один | 50 тыс. |
| Активно работающие участки | Более 90% генов | Менее 10% генов |
| Процессинг | Отсутствует | Осуществляется при переходе пре-мРНК из ядра в цитоплазму |
| Регуляция транскрипции | Оперонная | Сложная каскадная |
Как уже упоминалось выше, в отличие от прокариот основная часть генома эукариот находится в специальном клеточном компартменте (органелле), получившем название ядра , а значительно меньшая часть – в митохондриях, хлоропластах и других пластидах. Так же, как и у прокариот, информационной макромолекулой генома эукариот является ДНК, которая неравномерно распределена по нескольким хромосомам в виде комплексов с многочисленными белками. Эти ДНК-белковые комплексы эукариот получили название хроматина . На протяжении клеточного цикла хроматин претерпевает высокоупорядоченные структурные преобразования в виде последовательных конденсаций–деконденсаций. В соматических клетках при максимальной конденсации в метафазе митоза эти преобразования сопровождаются формированием видимых в микроскопе метафазных хромосом. Как морфология метафазных хромосом, так и их число являются уникальными характеристиками вида. Совокупность внешних признаков хромосомного набора эукариот получила название кариотипа . Эти признаки широко используются в биологической систематике.
Геном эукариот существенно отличается от генома прокариот по ряду признаков, среди которых необходимо отметить его избыточность. Содержание ДНК у эукариот в расчете на одну клетку в среднем на два–три порядка выше, чем у прокариот, и у разных видов животных изменяется от 168 пг (амфибии) до 1 пг (некоторые виды рыб). У человека имеется 6 пг ДНК на диплоидный геном, суммарная длина которой приближается к 6·10 9 п.о. (см. табл. I.1).
Повышенное содержание ДНК в геноме эукариот нельзя объяснить одним лишь увеличением потребности этих организмов в дополнительной генетической информации в связи с усложнением организации, поскольку большая часть их геномной ДНК, как правило, представлена некодирующими последовательностями нуклеотидов. Размер генома организмов, находящихся на более низких ступенях эволюционного развития, зачастую превышает размеры геномов более высокоорганизованных животных и растений. В настоящее время известно, что большая часть ДНК генома эукариот не кодирует РНК и белки, и ее генетические функции не вполне понятны. Особенности первичной структуры ДНК эукариот позволяют разделить ее на многочисленные семейства и классы, основные из которых кратко рассмотрены ниже.
Последовательности нуклеотидов эукариотического генома
Геном эукариот составляют уникальные и повторяющиеся последовательности нуклеотидов. Содержание уникальных последовательностей в геноме, определенное на основании кинетики реассоциации фрагментированной ДНК, варьирует у разных организмов, и их доля составляет 15–98% от всей ДНК. Несмотря на то, что во фракцию уникальных последовательностей попадают многие структурные гены, большая часть уникальных последовательностей является некодирующей и обычно не заключает в себе генетической информации в общепринятом значении этого термина: не кодирует функционально значимые полипептидные цепи или РНК. Хорошо известным примером таких уникальных последовательностей являются интроны, общий размер которых, как правило, на порядок и более превышает суммарный размер экзонов содержащих их генов.
Эволюционное возникновение мозаичной (интрон–экзонной) структуры генов эукариот, так же как и консервативный характер наследования размеров и взаимного расположения интронов в генах, не находит в настоящее время исчерпывающего объяснения из-за кажущегося отсутствия фактора давления естественного отбора на последовательности нуклеотидов без четких биологических функций. Наибольшее распространение получила концепция В. Гилберта (1977 г.), согласно которой появление интронов, по-видимому, совпавшее по времени с эволюционным возникновением многоклеточных организмов, обеспечило возможность обмена экзонами между неродственными генами (exon shuffling ). Такой обмен должен сопровождаться образованием новых белков мозаичного строения, составленных из готовых полипептидных функционально значимых модулей (доменов), ранее принадлежавших другим белкам. Следствием этого, по мнению сторонников данной концепции, было резкое ускорение образования белков и ферментов с новыми функциями, а также глубокие эволюционные преобразования самих организмов, реализующих такие молекулярные механизмы. Эта точка зрения получила название "гипотезы позднего возникновения интронов" (intron late ) . В соответствии с другой гипотезой Дж.Е. Дарнелла и В.Ф. Дулиттла (1978 г.) современные интроны представляют собой "эволюционные реликты". Когда-то интроны были частью гигантских генов.
Не менее загадочным с эволюционной точки зрения остается и феномен появления в геноме многоклеточных организмов большого количества некодирующих повторяющихся последовательностей. Такие повторы представлены в гаплоидном геноме эукариот множественными копиями. В современной классификации повторов принято различать часто повторяющиеся последовательности, число которых превышает 10 5 на гаплоидный геном, и умеренно повторяющиеся, представленные 10–10 4 копиями. Хорошо изученным представителем первых является сателлитная ДНК , которая состоит из коротких тандемных повторов длиной 1–20 п.о., организованных в длинные блоки. Одними из первых среди повторяющихся последовательностей ДНК эукариот были открыты сателлитные ДНК тимуса телят. Свое название они получили на основании того, что при анализе суммарной эукариотической ДНК центрифугированием в градиенте плотности хлористого цезия они сопровождали основной пик оптической плотности в виде плеча (спутника, сателлита). Именно гомогенный нуклеотидный состав фракции сателлитных ДНК, определяемый наличием в ней многочисленных коротких повторов, изменял ее плавучую плотность, что легко обнаруживалось при центрифугировании. В своем классическом определении сателлитных ДНК Р.Д. Бриттен и соавт. (1974 г.) отмечали, что сателлиты – это минорный компонент ДНК, отделяющийся от основной ДНК при равновесном ультрацентрифугировании в градиенте плотности CsCl . Для сателлитов характерен ряд свойств, среди которых наиболее важны: а) быстрая и точная реассоциация в процессе ренатурации ДНК; б) множество копий; в) простая первичная структура; г) гомогенный состав (протяженные кластеры одних и тех же повторяющихся блоков последовательны); д) пурин–пиримидиновая асимметрия в распределении нуклеотидов по цепям ДНК; е) концентрирование в прицентромерном гетерохроматине; ж) ограниченная репликация (недорипликация) при политенизации хромосом; з) нахождение в составе хромосом в виде тандемно (друг за другом) расположенных кластеров. Содержание сателлитной ДНК в геноме эукариот может достигать 5–50% от суммарного количества ДНК. Микро - (от 1 до 4 п.о. в основном повторяющемся блоке) и минисателлитные (с бó льшим числом п.о. в индивидуальном повторе) ДНК характеризуются высокой вариабельностью по числу копий в геномах организмов даже одного вида и в ряде случаев обладают генетической нестабильностью как в норме, так и при некоторых патологических состояниях организмов. Благодаря этому свойству мини- и микросателлиты часто называют тандемными повторами с изменяющимся числом копий VNTR (variable number of tandem repeats ).
Другой тип повторов – диспергированные повторяющиеся последовательности ДНК , не организованные в крупные блоки, а рассеянные по геному. Повторы этого типа, иначе называемые умеренно повторяющимися последовательностями (medium reiterated frequency repeats – MERs), разделяют на два обширных класса: SINE (short interspersed elements) – короткие и LINE (long interspersed elements) – длинные диспергированные элементы. Длина SINE-элементов составляет 90–400 п.о., тогда как длина LINE-последовательностей может достигать 7 т.п.о. Хорошо изученными повторами класса SINE в геноме человека и некоторых приматов являются так называемые Alu-повторы , длина повторяющейся единицы которых составляет 300 п.о. Alu-повторы представлены в геноме человека ~10 6 копиями и в среднем встречаются через каждые 4 т.п.о., составляя ~5% от суммарного количества ДНК. Аналогичные в структурном отношении повторы, названные B1, обнаружены в геноме мышей и под другими названиями описаны у многих млекопитающих.
Хотя LINE-последовательности заключают в себе гены обратных транскриптаз, что является признаком ретротранспозонов (мобильных генетических элементов животных, обладающих структурным сходством с геномом ретровирусов), для них характерно отсутствие последовательностей длинных концевых повторов (long terminal repeats – LTR), типичных для ретротранспозонов (подробнее о геноме ретровирусов см. раздел 7.2.7). В качестве примера LINE-последовательности можно упомянуть LINE-1-повтор, широко распространенный в геноме животных. LINE-1-элемент мышей содержит две открытые рамки считывания ORF-1 и ORF-2, вторая из которых кодирует белок, гомологичный обратной транскриптазе. ORF фланкированы короткими нетранслируемыми последовательностями, а сами LINE-1 – короткими прямыми повторами (SDR). 5’-Концевые последовательности повтора функционируют в качестве промоторов транскрипции. Этот участок LINE-1 грызунов (но не человека) построен из коротких тандемных повторов двух типов A и F, называемых мономерами. Длина мономеров у крыс составляет 600 п.о. При этом A- (но не F) мономеры обладают активностью промоторов.
Так же как и сателлитные ДНК, SINE- и LINE-повторы характеризуются генетической нестабильностью. Их общими чертами являются транскрибируемость и способность к транспозициям. Последовательности РНК, транскрибированные с умеренных повторов, обнаруживают среди гетерогенных ядерных РНК, где их доля достигает 20–30%. Имеются экспериментальные свидетельства того, что новые копии повторяющихся элементов обоих типов возникают в геноме в результате функционирования механизма, названного ретротранспозицией , или ретропозицией . При участии подобного механизма под действием обратной транскриптазы сначала образуется кДНК на матрице РНК-транскрипта соответствующего повтора, которая далее интегрируется в новый локус генома, как это имеет место у ретровирусов. Такой механизм дает возможность локально изменять число копий определенных последовательностей нуклеотидов в эукариотическом геноме. Тем не менее, большая часть LINE-последовательностей неспособна к транспозициям, и их ORF, по-видимому, могут быть отнесены к псевдогенам – неэкспрессирующимся последовательностям, гомологичным последовательностям истинных генов. Помимо вышеупомянутых повторяющихся последовательностей геном человека содержит более 100 000 копий MaLR-повторов длиной в 2–3 т.п.о., содержащих LTR, и несколько тысяч последовательностей генома ретровирусов.
Функциональную значимость избыточной ДНК лишь частично объясняют концепции, приписывающие ей структурную роль в пространственной организации генома и участие в конъюгации гомологичных хромосом в мейозе или репликации теломерных участков хромосом. Таким образом, основные положения знаменитого парадокса C, указывающие на необъяснимое присутствие в геноме эукариотических организмов большого количества избыточной ДНК, по-прежнему остаются загадочными и парадоксальными. Попытка нового объяснения основной функциональной роли "избыточной" ДНК в геноме эукариот сделана в разделе 5.3.
информации и ее регуляция
Цель. Знать молекулярную структуру и свойства нуклеиновых кислот, хромосом, стадии биосинтеза белка, принципы регуляции генной активности. Уметь выявлять ДНК в ядрах клеток с помощью реакции Фельгена.
Задание для студентов
Работа 1. ДНК в ядрах клеток
На постоянном препарате под большим увеличением микроскопа рассмотрите в ядрах клеток эпителия слизистой оболочки ротовой полости ДНК, выявленную с помощью реакции Фельгена.
Зарисуйте несколько ядер, в которых ДНК окрашена в пурпурномалиновый цвет.
Работа 2. Молекулярная структура ДНК эукариот
Рассмотрите рис. 1. Зарисуйте строение вторичной (2) структуры
Рис. 1. Строение ДНК эукариот.
Структуры ДНК: 1 - первичная; 2 - вторичная; 3 - третичная.
А - аденин; Г - гуанин - пуриновые азотистые основания; Ц - цитозин; Т - тимин-пиримидиновые азотистые основания; Д - дезоксирибоза; Ф - остаток фосфорной кислоты; Н - нуклеотид
Работа 3. Структурно-функциональная организация ДНК у про- и эукариот
Изучите таблицы, перепишите их в рабочую тетрадь.
Признаки | Прокариоты | Эукариоты |
Количество генов | 4 тыс. (Е. coli) | Около 30 тыс. (человек) |
Количество ДНК | 4 млн пар нуклеотидов | 3-7 млрд пар нуклеотидов |
Кодирующие последовательности | ||
Связь ДНК с гистонами | Отсутствует | Формирует нуклеосомы |
Укладка ДНК | Кольцевая, содержит 100 петель по 40 тыс. пар нуклеотидов | Линейная с замкнутыми в теломеры концами, имеет 4 уровня спирализации |
Количество репликонов | ||
Активно работающие участки | Более 90% генов | Менее 10% генов |
Процессинг | Отсутствует | Осуществляется при переходе пре-мРНК из ядра в цитоплазму |
Регуляция транскрипции | Оперонная | Сложная каскадная |
Работа 4. Организация наследственного материала у прокариот (нуклеоид)
Рассмотрите рис. 2 и обратите внимание на укладку ДНК в виде петель.
 Рис. 2.
Укладка ДНК в нуклеоиде прокариот:
Рис. 2.
Укладка ДНК в нуклеоиде прокариот:
1 - кольцевая молекула ДНК; 2 - укладка ДНК в виде петель; 3 - белки, связывающие петли ДНК
Работа 5. Уровни организации интерфазного хроматина
Рассмотрите по рис. 3 уровни организации наследственного материала у эукариот.
 Рис. 3.
Схема
различных уровней компактизации хроматина: а - нуклеосомная нить; б -
микрофибрилла; в - интерфазная хромонема; г - молекулярная организация
нуклеосомной нити: 1 - нуклеосома; 2 - ДНК; 3 - гистоны Н2А, Н2В, Н3 и
Н4; 4 - гистон Н1
Рис. 3.
Схема
различных уровней компактизации хроматина: а - нуклеосомная нить; б -
микрофибрилла; в - интерфазная хромонема; г - молекулярная организация
нуклеосомной нити: 1 - нуклеосома; 2 - ДНК; 3 - гистоны Н2А, Н2В, Н3 и
Н4; 4 - гистон Н1
Работа 6. Биосинтез белка у прокариот и эукариот
Изучите и зарисуйте процесс биосинтеза белка по схеме 1.
 Схема 1.
Биосинтез белка у прокариот (а) и эукариот (б)
Схема 1.
Биосинтез белка у прокариот (а) и эукариот (б)
Работа 7. Транскрипция и процессинг у эукариот
Изучите транскрипцию и процессинг по рис. 4.
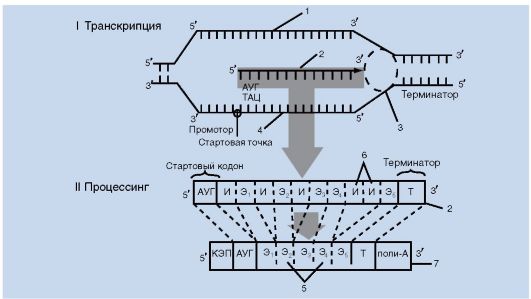 Рис. 4.
Транскрипция и процессинг у эукариот:
Рис. 4.
Транскрипция и процессинг у эукариот:
1 - ДНК; 2 - пре-мРНК; 3 - РНК-полимераза; 4 - кодогенная цепь ДНК; 5 - экзоны; 6 - интроны; 7 - зрелая мРНК; Т - терминатор; КЭП и поли-А - концевые последовательности нуклеотидов; ТАЦ и АУГ - инициаторные триплеты
Работа 8. Трансляция. Этапы рибосомного цикла
Изучите и зарисуйте по рис. 5 процесс трансляции.
 Рис. 5.
Процесс трансляции:
Рис. 5.
Процесс трансляции:
1 - малая субъединица рибосомы; 2 - большая субъединица рибосомы; 3 - аминоацильный (А) центр; 4 - пептидильный (П) центр; 5 - АУГ-инициаторный триплет мРНК; 6 - терминатор мРНК; 7 - инициаторная тРНК; 8 - аминокислоты формирующегося полипептида; 9 - колпачок
Работа 9. Регуляция активности генов у прокариот (схема Жакоба-Моно)
Рассмотрите и зарисуйте изображение регуляции синтеза белка путем индукции и репрессии (рис. 6).

 Рис. 6.
Регуляция
синтеза белка путем индукции (а, б) и репрессии (в, г): а - структурные
гены оперона блокированы; б - дерепрессирование генов индуктором; в -
при недостаточном количестве конечного продукта (корепрессора) оперон
дерепрессирован, а при избыточном - блокирован (г)
Рис. 6.
Регуляция
синтеза белка путем индукции (а, б) и репрессии (в, г): а - структурные
гены оперона блокированы; б - дерепрессирование генов индуктором; в -
при недостаточном количестве конечного продукта (корепрессора) оперон
дерепрессирован, а при избыточном - блокирован (г)
Работа 10. Основные принципы регуляции активности генов у эукариот
Изучите и перепишите.
1. У эукариот не установлено оперонной организации генов, так как гены, определяющие синтез ферментов одной цепи биохимических реакций, могут быть рассеяны в геноме и не имеют, как у прокариот, единой регулирующей системы (ген-регулятор, промотор, оператор и т.д.).
2. Регуляция транскрипции у эукариот комбинационная, т.е. активность каждого гена регулируется большим числом генов-регуляторов.
3. У многих эукариотических генов в ДНК имеется несколько зон, узнаваемых разными белками.
4. У эукариот существуют белки-регуляторы, контролирующие работу других регуляторных белков, и их действие может характеризоваться плейотропным эффектом.
5. В регуляции экспрессии эукариотических генов важную роль играют гены энхансеры (усиливают транскрипцию) и сайленсеры (тормозят транскрипцию).
6. В регуляции транскрипции участвуют гормоны, а генной активности - гистоны хромосом.
7. Регуляция экспрессии генов осуществляется на всех этапах реализации наследственной информации.
Вопросы для самоподготовки
1. Каковы особенности организации наследственного материала у про- и эукариот?
2. Какова молекулярная организация и функции нуклеиновых кислот?
3. Что такое ген? Какое определение гена Вы считаете более точным?
4. Каковы особенности строения генов у про- и эукариот?
5. Что такое генетический код и каковы его свойства?
6. Каковы основные этапы биосинтеза белка, в чем их сущность?
7. Каковы механизмы регуляции генной активности у прокариот (схема Жакоба-Моно)?
8. Каковы основные принципы регуляции генной активности у эукариот?
Тестовые задания
Выберите один правильный ответ.
1. ЭЛЕМЕНТАРНОЙ ЕДИНИЦЕЙ ФУНКЦИИ НАСЛЕДСТВЕННОГО МАТЕРИАЛА ЯВЛЯЕТСЯ:
2. ТРАНСКРИПЦИЮ ОСУЩЕСТВЛЯЕТ ФЕРМЕНТ
1. ДНК-полимераза
2. РНК-полимераза
3. Геликаза
3. МУЛЬТИГЕННЫЕ СЕМЕЙСТВА И КОМПЛЕКСЫ В ГЕНОМЕ
1. Прокариоты
3. Эукариоты
Выберите несколько правильных ответов.
4. СВОЙСТВАМИ ДНК КАК ВЕЩЕСТВА НАСЛЕДСТВЕННОСТИ ЯВЛЯЮТСЯ:
1. Химическая стабильность
2. Репликация
3. Репарация
4. Способность к трансляции
5. БИОСИНТЕЗ БЕЛКА ПРОИСХОДИТ С УЧАСТИЕМ ОРГАНЕЛЛ:
1. Лизосомы
2. Гладкая ЭПС
3. Рибосомы
4. Полисомы
6. ОСОБЕННОСТЯМИ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНОВ У ЭУКАРИОТ ЯВЛЯЮТСЯ:
1. Отсутствие оперонной организации генов
2. Наличие оперонной организации генов
3. Наличие комбинационной регуляции транскрипции
4. Регуляция экспрессии генов на всех этапах реализации генетической информации
Установите соответствие.
7. ТРИПЛЕТЫ ДНК:
ТРИПЛЕТЫ мРНК:
Установите правильную последовательность.
8. УПАКОВКИ ДНК В ХРОМОСОМЕ ЭУКАРИОТ:
1. Хромонема
2. Хроматида
3. Нуклеосомная нить
4. Микрофибрилла
9. БИОСИНТЕЗА БЕЛКА У ЭУКАРИОТ:
1. Трансляция
2. Транскрипция
3. Процессинг
4. Посттрансляция
10. РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНОВ У ПРОКАРИОТ
(СХЕМА ЖАКОБА-МОНО):
1. Считывание информации со структурных генов
2. Образование комплекса индуктор-репрессор
3. Поступление индуктора в цитоплазму прокариота
4. Освобождение оператора от репрессора
5. Образование полицистронного транскрипта
6. Синтез отдельных пептидов
Литература
Основная
Биология / Под ред. В.Н. Ярыгина. - М.: Высшая школа, 2001. - Кн. 1. - С. 65-138, 147-152, 163-171.
Пехов А.П. Биология с общей генетикой. - М.: Изд-во РУДН, 1993. - С. 95-112, 141-154, 166-171.
Дополнительная
Альберт Б. и др. Молекулярная биология клетки. - М.: Мир, 1994. -
Гильберт С. Биология развития. - М.: Мир, 1994. Жимулев И.Ф. Общая и молекулярная генетика. - Н.: Сибирское университетское издательство, 2003.
Длина ДНК в ядре намного больше, чем размер камеры, в которой он содержится. Например, самая короткая хромосомы человека содержит 4,6 х 107 п.н. ДНК (примерно в 10 раз больше размера генома E.coli). Это эквивалентно 14000 мкм расширенного ДНК. В наиболее конденсированном состоянии во время митоза, хромосома около 2 мкм длиной. Это дает соотношение упаковки 7000 (14000/2).
Для достижения общего коэффициента упаковки ДНК не упакован непосредственно в окончательную структуру хроматина. Вместо этого он содержит несколько иерархий организации. Первый уровень упаковки достигается путем намотки ДНК вокруг белкового ядра для получения шарообразной структуры, называемой нуклеосома. Это дает упаковке соотношение около 6. Эта структура инвариантна как в эухроматине и гетерохроматине всех хромосом.
Второй уровень упаковки является намоткой из бисера в спиральную структуру называемую 30 нм волокно, которое находится в межфазной хроматине и митотической хромосоме. Эта структура увеличивает коэффициент упаковки до 40.
Окончательная упаковки происходит, когда волокно организовано в петли, при этом конечный коэффициент упаковки равен около 1000 в интерфазе хромосом и около 10000 в митотических хромосомах.
Эукариотические хромосомы состоят из ДНК-белкового комплекса, который организован в компактной форме, и она позволяет хранить большое количество ДНК, которые будут сохранены в ядро клетки. Субъединица, обозначающая хромосомы, называется хроматин. Основной единицей хроматина является нуклеосом.
Хроматина – единица анализа хромосом; хроматина отражает общую структуру в хромосоме, но не является уникальной для какой-либо конкретной хромосомы.
Нуклеосом – простая упаковка структуры ДНК, которая находится во всех эукариотических хромосомах; ДНК обернута вокруг малых основных белков, называемых гистонами.
Жизнь эукариотической клетки
Эукариоты имеют оформленное ядро, содержащее ДНК. Размер типичной эукариотической клетки, например, клетки печени человека, составляет в поперечнике ~ 25 мкм. Ее ядро, размером ~ 5 мкм в диаметре, содержит 46 хромосом, суммарная длина ДНК которых равна 2 м. Эукариоты содержат значительно больше ДНК, чем прокариоты. Так, клетки человека и других млекопитающих содержат в 600 раз больше ДНК, чем в Е.coli. Общая длина всей ДНК, выделенной из клеток организма взрослого человека, составляет ~ 2 х 10 13 м или 2 х 10 10 км, что превышает окружность земного шара (4 х 10 4 км) и расстояние от Земли до Солнца (1,44 х 10 8 км).
У эукариот ДНК находится в хромосомах. В клетках человека …
46 хромосом (хроматид), которые организованы в 23 пары. Каждая хромосома эукариотической клетки содержит одну очень большую молекулу двухцепочечной ДНК, несущую набор генов. Совокупность генов клетки составляет ее геном.
Гены
– это участки ДНК, которые кодируют полипептидные цепи и РНК.
Молекулы ДНК в 46 хромосомах человека не одинаковы по размеру. Средняя длина хромосомы составляет 130 млн. пар оснований и имеет длину 5 см. Понятно, что уместить такой длины ДНК в ядре возможно только путем ее определенной упаковки. При образовании третичной структуры ДНК человека происходит в среднем уменьшение ее размеров в 100 тысяч раз.
Упаковка ДНК в эукариотических хромосомах отличается от ее упаковки в прокариотических хромосомах. Эукариотические ДНК имеют не кольцевую, а линейную двухцепочечную структуру. Кроме того, третичная структура ДНК у эукариотических клеток отличается тем, что многократная спирализация ДНК сопровождается образованием комплексов с белками. ДНК эукариот содержит экзоны — участки, кодирующие полипептидные цепи, и интроны – некодирующие участки (выполняют регуляторную функцию).
Эукариотические хромосомы состоят из хроматиновых волокон .
Эукариотические хромосомы выглядят как резко очерченные структуры только непосредственно до и во время митоза- процесса деления ядра в соматических клетках. В покоящихся, неделящихся эукариотических клетках хромосомный материал, называемый хроматином , выглядит нечетко и как бы беспорядочно распределен по всему ядру. Однако, когда клетка готовится к делению, хроматин уплотняется и собирается в хромосомы.
Хроматин состоит из очень тонких волокон, которые содержат ~ 60 % белка, ~ 35 % ДНК, и, вероятно, ~ 5 % РНК. Хроматиновые волокна в хромосоме свернуты и образуют множество узелков и петель. ДНК в хроматине прочно связана с белками-гистонами, функция которых состоит в упаковке и упорядочении ДНК в структурные единицы – нуклеосомы. В хроматине содержится также ряд негистоновых белков. Хроматиновые волокна напоминают по внешнему виду нитки бус. Бусинки – это нуклеосомы .
Нуклеосома состоит из белков-гистонов. Каждая нуклеосома содержит 8 молекул гистонов – по 2 молекулы Н2А. Н2В, Н3, Н4. Двухцепочечная ДНК обвивает нуклеосому дважды.
Нить ДНК намотана на гистоновое ядро нуклеосомы снаружи. В промежутках между нуклеосомами расположена соединительная нить ДНК, с которой связывается гистон Н1. Таким образом, нуклеосомы – это структурные единицы хроматина, выполняют функцию плотной упаковки ДНК. (ДНК укорачивается за счет того, что она обвивает гистоны). Хроматин связан также с негистоновыми белками ядра, которые образуют ядерный матрикс.

Эукариотические клетки содержат также цитоплазматическую ДНК .
Кроме ДНК в ядре у эукариот есть ДНК в митохондриях . Хлоропласты фотосинтезирующих клеток также содержат ДНК. Обычно ДНК в цитоплазме составляет » 0, 1 % всей клеточной ДНК.
Митохондриальные ДНК – это двухцепочечные кольцевые молекулы малого размера.
Молекулы ДНК в хлоропластах значительно больше, чем в митохондриях.
ДНК митохондрий и хлоропластов не связана с гистонами.